
ARTÍCULO ORIGINAL
Biotecnología
Vegetal Vol. 14, No. 2: 107 - 115, abril - junio, 2014
ISSN 2074-8647, RNPS: 2154 (Versión electrónica)
Instituto de Biotecnología de las Plantas. UCLV. MES.
Efecto de diferentes concentraciones de cobre en el medio de cultivo sobre plantas in vitro de Agave fourcroydes L.
Effect of different cupper concentration in culture media on Agave fourcroydes L. in vitro plants
Mohamed Sankoumba Fadiga, Cúc Ðoàn ThË Thu, Yunel Pérez, Gerardo González*. *Autor para correspondencia
Facultad de Agronomía, Universidad de Matanzas Camilo Cienfuegos. Autopista a Varadero km 3½. Matanzas. Cuba. e-mail: oramascuba07@gmail.com.
RESUMEN
Se realizó una investigación para determinar el efecto del cobre sobre plantas in vitro de henequén (Agave fourcroydes L.) var. `Sac Ki'. Se adicionó CuSO4 (25, 50, 75, 100 µM) en el medio de cultivo. Después de 30 días de establecidas las plantas in vitro se determinó la producción de biomasa, el contenido de carbohidratos solubles totales, azúcares reductores, proteínas solubles, la actividad específica de las enzimas catalasa y peroxidasa y el número de brotes. No se observaron anormalidades fenotípicas o síntomas de toxicidad en las plantas. La biomasa se incrementó a la concentración de 50 µM y el número de brotes a 25 µM. Se comprobó que el efecto sobre las variables evaluadas se relacionó con la concentración e indicaron tolerancia al cobre. Los resultados de este trabajo sientan las bases para el posible empleo de esta especie en la biorremediación de suelos contaminados.
Palabras clave: biorremediación, fitorremediación, henequén, metales pesados
ABSTRACT
A research was realized in order to determine the effect of copper on henequen (Agave fourcroydes L.) var. `Sac ki' in vitro plants. CuSO4 (25, 50, 75, 100 µM) was added to the culture medium. After 30 days of the in vitro plants established, the following indicators were determined: biomass production, soluble carbohydrates, reducing sugars, soluble proteins, specific activity of catalase and peroxidase enzymes and number of shoots. No phenotypic abnormalities or toxicity symptoms were observed in plants. Biomass showed an increase at the concentration of 50 µM also as the number of shoots at 25 µM. It was proven that the effect on the evaluated indicators was related to the concentration of copper and indicated the tolerance to copper. The results of this investigation settle down the bases for the possible use of this specie in bioremediation of contaminated soils.
Keywords: bioremediation, heavy metals, phytoremediation
INTRODUCCIÓN
Con el desarrollo de la sociedad humana, una gran cantidad de desechos y contaminantes se incorporan al medio ambiente. Entre ellos, se encuentran los metales pesados que se han convertido en una amenaza no sólo para el ecosistema, sino también para la salud humana, por la alta toxicidad y gran resistencia en el suelo. Estos pueden llegar al hombre a través de la cadena alimenticia o en las aguas (Barceló y Poschenrieder, 2003).
Para descontaminar
estos metales se han desarrollado métodos físicos, químicos
y biológicos. Entre estos últimos se encuentra el empleo de plantas
capaces de vivir en suelos
contaminados: la fitorremediación. Este método llama la atención
porque permite limpiar los suelos contaminados de una gran extensión,
además de ser internacionalmente reconocida como uno de los más
baratos, eficaces y respetuosos con el medio ambiente.
Agave fourcroydes L. (henequén) es un cultivo capaz de vivir en suelos pobres de nutrientes y salinos, es una planta micorrizada y además es productora de ácidos orgánicos (Nobel, 2009). Estas características la hacen prometedora para ser utilizada como fitorremediadora de suelos contaminados con metales pesados. Además, cabe resaltar que el henequén al no entrar en la cadena alimentaria aparta los riegos de afectación al ser humano con la inmovilización de los metales pesados y cumple el doble propósito de producción industrial y fitorremediación.
Los estudios sobre la respuesta de henequén en condiciones de estrés abiótico, específicamente, de suelos contaminados, son escasos, y por ello requieren más atención e investigación para afirmar su capacidad como planta fitorremediadora.
Aunque no existen trabajos que indiquen el efecto de los altos niveles de metales pesados en el suelo sobre el desarrollo de las plantas de henequén, esta especie crece naturalmente sobre suelo de serpentinita el cual posee elevadas concentraciones de Ni y Cr (Chiarucci y Baker, 2007).
Existen especies de Agave que pueden crecer en suelos con altas concentraciones de metales pesados sin daños severos. Por ejemplo, Agave sisalana Perr., estrechamente relacionada con A. fourcroydes, fue clasificada como hiperacumuladora de Cr y como bioindicator de Zn, Pb y Ni (Mganga et al., 2011).
Por lo antes señalado, este trabajo se propuso como objetivo determinar el efecto de la adición de cobre (CuSO4) en el medio de cultivo sobre plantas de henequén establecidas in vitro.
MATERIALES Y MÉTODOS
Material vegetal
Plantas seleccionadas en campo de la variedad `Sac Ki' o henequén blanco, procedentes del germoplasma de la Empresa Henequenera de Matanzas Eladio Hernández, se utilizaron como fuente de explantes para el establecimiento in vitro.
Las plantas se lavaron con agua y detergente (1.0% m/v) por 30 minutos. Se extrajeron ápices de 7 cm de longitud que se desinfectaron superficialmente por inmersión en solución de alcohol al 70% (v/v) durante un minuto. A continuación se flamearon hasta extinción del fuego. Seguidamente se introdujeron en solución de hipoclorito de sodio 2.5% (v/v) durante 20 minutos y posteriormente se realizaron tres enjuagues, durante 25 minutos (5 10 10 minutos) con agua destilada estéril. Finalmente, con la ayuda de un cuchillo se eliminaron las partes del material vegetal dañado.
Ya en la cabina de flujo laminar se redujo su tamaño a 4.0 cm de longitud y se sumergieron en solución de hipoclorito de sodio al 2.5% (v/v) durante 10 minutos. Se realizaron tres enjuagues, durante 15 minutos (5 5 5 minutos) con agua destilada estéril y se eliminaron las partes del material vegetal dañadas y se redujo su longitud a 1.5 - 2.0 cm.
Los explantes se colocaron en medio de cultivo MS, 0.1 mg l-1 de tiamina, 0.5 mg l-1 de piridoxina; 0.5 mg l-1 de ácido nicotínico, 2.0 mg l-1 de glicina, 100 mg l-1 de mio-inositol y 30 g l-1 de sacarosa. Se adicionaron 0.025 mg l-1 de ácido 2,4 diclorofenoxiacético (2,4-D) y 10.0 mg l-1 de 6-bencilaminopurina (6-BAP). El medio de cultivo fue solidificado con 10 g l-1 de Agar Técnico Agar #3 (Biocen). El pH fue ajustado a 5.7 antes de la esterilización a 121°C y 1.2 kg cm-2. En frascos de vidrio de 250 ml de capacidad se añadieron 30 ml de medio de cultivo. Se colocaron cuatro explantes por frasco y se mantuvieron en cámaras de crecimiento con luz solar a 26 ± 2ºC (37.5 µmol m2 s-1 de luminosidad como promedio).
Después de una semana de cultivo los explantes se transfirieron a frascos de cultivo con similar medio de cultivo y al que se añadieron diferentes concentraciones de CuSO4. 5H20 (25, 50, 75, 100 µM) con las cuales se conformaron cuatro tratamientos. Además, se incluyó como control el medio de cultivo libre de CuSO4. Se colocaron cinco plantas por frasco y el número de frascos por tratamiento fue de 10. Estos se mantuvieron durante 30 días en similares condiciones a las descritas anteriormente. Al final del periodo se realizaron las evaluaciones que se mencionan más abajo.
Contenido de biomasa
Con el propósito de valorar el efecto del CuSO4 en la acumulación de biomasa de las plantas in vitro se tomaron cinco plantas por tratamiento y se secaron en estufa a 60ºC por 72 horas, hasta peso constante para determinar la masa seca (g).
Contenido de proteínas solubles
Se tomaron muestras foliares (hoja más desarrollada) (0.5 g) de cinco explantes por tratamiento y se trituraron en 2.0 ml de solución tampón de fosfato de sodio 50 mM, (pH 7.0) en frío, la muestra fue centrifugada a 10 000 rpm durante 20 minutos a 4oC y se extrajo el sobrenadante para la medición inmediata del contenido de proteínas solubles. La concentración de proteínas solubles fue medida por el método de Bradford (1976).
Actividad de enzimas catalasa y peroxidasa
Se tomaron muestras foliares (hoja más desarrollada) (0.5 g) de tres explantes por tratamiento y se trituraron en 2 ml de solución tampón de fosfato de sodio 50 mM (pH 7.0) en frío, la muestra fue centrifugada a 10 000 rpm durante 20 minutos a 4oC. Se extrajo el sobrenadante para la medición inmediata de la actividad enzimática de catalasa y peroxidasa.
La actividad de catalasa fue determinada por medición de la velocidad de la descomposición del H2O2 previamente ajustada a una DO240 (densidad óptica de 240nm) (0.040-0.045) utilizando como blanco agua destilada, en una solución tampón de fosfato de potasio 20 mM, pH 7.0 que contenía 3% (v/v) de H2O2. El volumen final del ensayo fue de 3 ml. La lectura fue realizada a 240 nm en un espectrofotómetro (Ultrospect 2000).
Se tomaron tres mediciones por muestra en el espectrofotómetro (Ultrospect 2000). La actividad enzimática se calculó según la fórmula:

Contenido de carbohidratos solubles totales y azúcares reductores
Se determinó el contenido de azúcares reductores y carbohidratos solubles totales a partir de la extracción realizada para la evaluación de la actividad enzimática. Se siguió la metodología modificada del FenolSulfúrico para determinar el contenido de carbohidratos totales (Mahadevan y Sridhar, 1986). Los azúcares reductores se determinaron según la técnica de Dinitrosalisílico (Sumner, 1921). Para ambas determinaciones se establecieron curvas patrones (y= 5.3909x 0.0194. R2 = 0.9796 para azúcares reductores: y= 4.6241x 0.0692. R2 = 0.999 para carbohidratos solubles totales).
Efecto sobre el número de brotes
A los 30 días de cultivo en los diferentes tratamientos se cuantificó el número de brotes por planta con el objetivo de valorar el efecto del Cu en la multiplicación in vitro.
Análisis estadístico
Los experimentos se realizaron según un diseño completamente aleatorizado con tres réplicas para cada tratamiento. Los datos se procesaron mediante el paquete Statgraphic Versión 5.0 (2000), sobre WINDOW. Se determinó si los datos tenían Distribución Normal mediante la prueba de Bondad de Ajuste Kolmogorov - Smirnov y Homogeneidad de Varianza mediante las Pruebas de Bartlett. En los casos en que los datos cumplieron los supuestos se procesaron mediante ANOVA de clasificación simple y las medias se compararon mediante la Prueba de Rangos Múltiples de Duncan.
RESULTADOS Y DISCUSIÓN
Contenido de biomasa
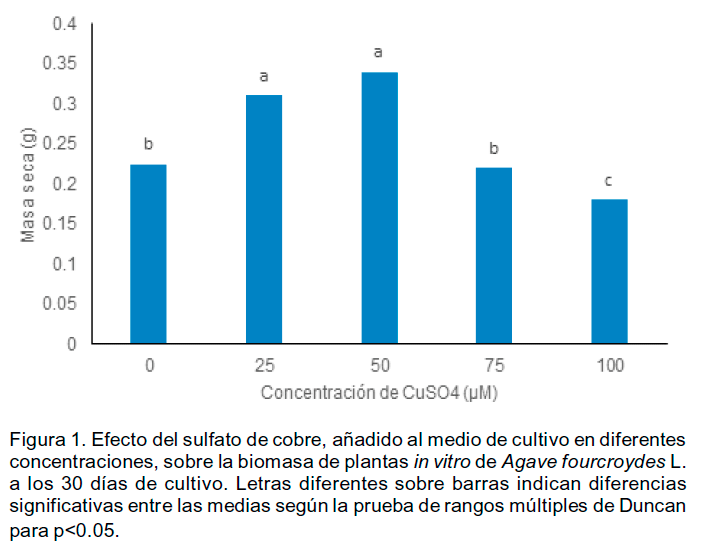
Se comprobó que la inclusión de CuSO4 en el medio de cultivo, excepto a la concentraciones de 25, 50 y 100 µM, no afectó el contenido de masa seca de las plantas in vitro (Figura 1).
La actividad de peroxidasa se midió por el método guayacol peroxidasa (Bergmeyer, 1974). El ensayo de actividad enzimática se realizó en un volumen final de 3 ml. Para la reacción se adicionó en una cubeta de cuarzo 2.80 ml de solución tampón fosfato de sodio 0.1 M, pH 7.0, 50 µl de solución guayacol 0.018 M, 50 µl solución de peróxido de hidrógeno ajustado a DO240 (0.040-0.045) utilizando agua como blanco. A la mezcla reaccionante se adicionaron 100 µl del extracto vegetal. La actividad enzimática se realizó a 25ºC y se midió a DO436. Se tomaron tres mediciones por muestra en un Espectrofotómetro (Ultrospect 2000). La actividad enzimática se calculó según la fórmula:
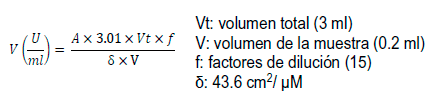
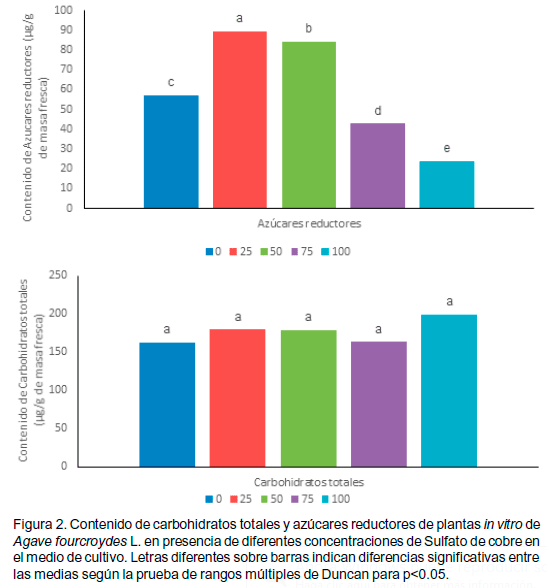
ensayados. Sin embargo, para la variable azúcares reductores, los tratamientos se diferenciaron significativamente del control (Figura 2). El mayor valor se encontró a la concentración de 25µM de cobre y el menor valor a 100µM.
En relación con el contenido de carbohidratos a pesar de la presencia de diferentes concentraciones de cobre en el medio de cultivo, se mantuvo similar entre todos los tratamientos, lo que indica la no alteración de la síntesis de estas sustancias.
La producción de carbohidratos se relaciona con una adecuada concentración de cobre. En este sentido, Mohamed (1986) encontró una relación entre cobre y ferredoxil en el sitio reductor de PSI donde Cu2+ estimuló el conjunto de transporte de electrón desde agua hacia NADP y se produjo mayor síntesis de carbohidratos. Según Deff (2007), en presencia de Cu se estimulan los fotosistemas I y II de la fotosíntesis, lo que sugiere en este caso la mixotrofía del cultivo.
El aumento de contenido de carbohidratos también se estimula a bajas concentraciones de cobre, tal es el caso del cultivo de Rosmarinus officilalis L (Deff, 2007) o con otros metales pesados en el cultivo de Lupinus albus (Costa y Spitz, 1997).
Los resultados indicaron un incremento significativo de la biomasa de las plantas in vitro con las concentraciones de 50µM y 25µM). Este comportamiento se diferencia de otros cultivos. De acuerdo con lo planteado por Costa y Spriz (1997) la tendencia de aumentar la masa seca al aumentar la concentración de Cu hasta cierto nivel puede ser debido a la acumulación de metal y esta no afecta a la asimilación normal de CO2.
Cu es un micronutriente esencial que participa en múltiples procesos metabólicos y en las concentraciones adecuadas favorece el crecimiento de las plantas aunque en presencia de elevados niveles, normalmente estas presentan reducción de la biomasa y síntomas de clorosis (Yruela, 2005). Se ha informado que en exceso el cobre causa una alteración en la estructura de los cloroplastos. Sin embargo, en este estudio no se observó clorosis en las plantas evaluadas. La biomasa vegetal y en específico la producción de masa seca son indicadores para evaluar el crecimiento vegetal.
Contenido de azúcares reductores y carbohidratos totales
No se obtuvieron diferencias significativas, en cuanto al contenido de carbohidratos solubles totales entre los tratamientos
Por lo antes señalado no es contradictorio que en el control a pesar de no utilizarse altas concentraciones de cobre presentó un contenido similar de carbohidratos al del resto de los tratamientos pues en el medio de cultivo MS se agrega 0.10µM de CuSO4.5H20 (Golden Bell), valor inferior a los empleados en los tratamientos. Todo ello sugiere la plasticidad del henequén, para producir cantidades adecuadas de carbohidratos, bajo un rango amplio de concentraciones de cobre, que en este ensayo no fueron limitante.
Bajo las condiciones del cultivo in vitro el contenido de azúcares reductores en el control se toma como lo normal, por lo tanto un incremento o decremento de esta variable puede ser el resultado de cambios en el metabolismo celular.
Los resultados sugieren incremento de la respiración mitocondrial, menor actividad mixotrófica o mayor translocación para hojas más jóvenes o el tallo en las concentraciones de 75 y 100µM en relación con el control, sin embargo lo contrario se apreció en la concentraciones de 25 y 50 µM.
Deff (2007) planteó que el bajo contenido de azúcares reductores se atribuye al incremento de la respiración en los órganos de la planta. En relación con lo anterior, Ahmed et al. (2008) encontraron que en cultivo de Zea mays L. con tratamiento de metales pesados se incrementó la respiración y disminuyó la fotosíntesis.
El incremento o decremento del contenido de azúcares reductores podría indicar una mayor o menor actividad de enzimas hidrolíticas por la presencia de concentraciones diferenciales de Cu. Mohamed (1986) sugirió que la acumulación de azúcares reductores en hojas de plantas de tomate (Lycopersicon esculentum Mill.) se debió a la presencia de metales pesados como Mn, Zn y Fe, los que estimularon la actividad amilolítica y como resultado condujo a un incremento de azúcares solubles más que de polisacáridos.
Contenido de proteínas solubles
Las concentraciones de cobre empleadas favorecieron significativamente el incremento del contenido de proteínas solubles en relación con el control (Figura 3).

La presencia de cobre en altas concentraciones puede favorecer el contenido de proteínas dado la necesidad de éstas para desarrollar el proceso de respiración mitocondrial para el cual se establece el Ciclo de Krebs donde se producen los metabolitos necesarios para la producción de aminoácidos. Por otra parte, el crecimiento y desarrollo del vegetal requiere de proliferación y diferenciación celular lo cual necesita de proteínas. Por último, existen proteínas transportadoras de iones metálicos que actúan en el mantenimiento del balance iónico dentro de la célula (Kupper et al., 2009)
La tolerancia del henequén a la mayor concentración de Cu pudiera relacionarse con la función de este como activador de la biosíntesis de fitoquelatinas que es un péptido rico en cisteína y con alta capacidad de formación de complejos con los metales pesados, lo que le permite mantener un nivel de proteínas similar a la cuantificada en los restantes tratamientos, a pesar, de la reducción de la masa seca.
Es conocido el importante papel que desempeña el cobre en los complejos enzimáticos y reacciones redox que se desarrollan en las células lo que pudiera relacionarse con el incremento en la síntesis de proteínas. Según los resultados obtenidos, se observa que el control presentó el menor contenido de proteínas y el mayor incremento ocurrió con la adición de CuSO4. Esto sugiere que los niveles endógenos de cobre presentes en el control no ocasionaron el comportamiento de este como cuando se presenta en elevadas concentraciones, por ende no desencadenó los mecanismos de defensa de la planta contra este fenómeno, por lo tanto no se estimuló la síntesis de proteínas. Esto pudiera indicar que el cobre favoreció la síntesis de proteína una vez que se incrementó sus niveles en relación con la concentración que se emplea en la propagación in vitro del henequén.
Actividad de enzimas catalasa y peroxidasa
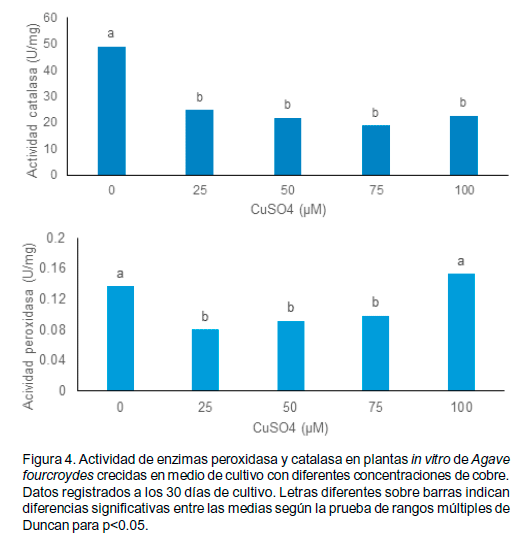
La actividad catalasa se inhibió en presencia de elevadas concentraciones de cobre, en el control se produjo el nivel de actividad específica de catalasa significativamente mayor (Figura 4).
Este resultado sugiere que la actividad de esta enzima se produce al inicio o primeros momentos del tratamiento con cobre y posterior a un tiempo disminuye. Esto se puede afirmar ya que no hubo signo de afectación fisiológica de los metales pesados sobre las plantas, señalando su actividad anterior, además de que el control con solo las concentraciones endógenas presentó una mayor actividad de esta enzima. Por otra parte, la respuesta positiva de las plantas junto con la baja actividad de la enzima catalasa pudiera estar relacionada con el eficiente funcionamiento del conjunto de mecanismos de protección de que dispone el henequén ante las elevadas concentraciones de cobre.
Betancourt et
al. (2005) plantearon que los metales pesados y el estrés abiótico
provocan cambios desfavorables en las plantas y basado en esto investigaciones
posteriores revelaron incrementos en la actividad enzimática de la catalasa
y la superóxido dismutasa en el citosol de raíces de Pisum
sativa contrario a lo sucedido en este ensayo lo que pudiera indicar
una relación entre los factores abióticos y la especie.
En el caso de la peroxidasa, la actividad no se diferenció entre los tratamientos de 25 µM, 50 µM y 75 µM. La actividad más elevada se registró en el control y el tratamiento de 100 µM.
Los metales pesados tales como el cobre, por sus propiedades, son capaces de estimular la formación de las especies reactivas de oxígeno, los cuales en alta concentración pueden dar lugar a una oxidación incontrolable de las estructuras de la célula, incluyendo ADN, proteínas y lípidos de membrana, lo que favorece la interrupción del metabolismo y la destrucción de la estructura celular además de que las plantas generan especies reactivas del oxígeno (Desikan et al., 2005). Por eso, no es sorprendente que ellas también poseen el sistema de defensa antioxidante integral que contiene componentes tanto enzimáticos como no enzimáticos, para controlar en niveles no tóxicos los radicales libres, manteniendo el balance redox celular.
De hecho, algunos de los procesos antioxidantes son inducibles respondiendo a la alta concentración de las ERO, lo que sugiere que los mecanismos celulares evolucionan para reconocer y mejorar su acción ante el alto contenido de tales compuestos (Desikan et al., 2005). Uno de ellos es la estimulación de la formación de las enzimas antioxidantes.
En este sentido, las evidencias acumuladas señalan que el cobre induce la actividad peroxidasa en plantas como por en frijol (Phaseolus vulgaris L. cv. Akman) o en ajo (Allium sativum) (Meng et al., 2007). Por otra parte, el cobre por ser un metal de transición cataliza la formación de H2O2, por lo tanto se supone que el aumento en concentración de esta sustancia tiene como consecuencia el aumento de la actividad peroxidasa. La respuesta obtenida sugiere además que en la concentración más elevada el mecanismo de secuestro ya no es suficiente para reducir la cantidad de zinc en el citoplasma por lo tanto se incrementa la formación de los ROS y estas últimas se convierten en la señal de activación de la peroxidasa (Desikan et al., 2005).
Efecto sobre el número de brotes
La diferenciación de plantas completas y el inicio de la multiplicación en los explantes de henequén se lograron en todos los tratamientos empleados. En presencia de 25 µM de CuSO4 (Figura 5) se superó significativamente el número de brotes en relación con el control.
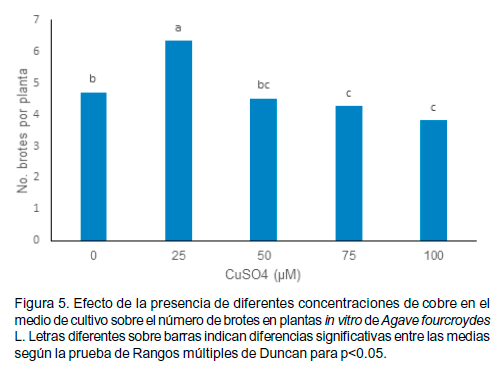
Ello sugiere que a esta concentración se estimuló la diferenciación de yemas axilares, lo que requiere de estudios posteriores con el objetivo de mejorar la micropropagación del henequén.
Las plantas producidas no mostraron anormalidades fenotípicas lo que sugiere que bajo estas condiciones (elevadas concentraciones de cobre) no se producen daños que pudieran ser letales y que interfieran con el crecimiento y desarrollo de este cultivo.
Se hace necesario tener en cuenta, que el Agave fourcroydes L. es una planta CAM en la cual, la producción de ácido orgánico (ácido málico) es muy elevada (Nobel, 2009). Este ácido posee la capacidad de reaccionar con los metales pesados a través de los enlaces débiles de oxígeno. Küpper et al. (2009) plantearon que este tipo de enlace es esencial en la tolerancia de las plantas hiperacumuladoras de metales pesados.
En las plantas hiperacumuladoras de metales pesados se produce la reacción de casi todos estos con ácidos orgánicos, los cuales se encuentran comúnmente en vacuola. Además de los posibles mecanismos de tolerancia a los metales pesados mencionados durante toda la discusión del trabajo, el Agave fourcroydes L. posee este recurso que evidencia la estabilidad de su metabolismo a altas concentraciones cobre, y por lo tanto, podría significar uno de los mecanismo que justifican la tolerancia a metales pesados.
Estudios futuros acerca de la tolerancia del A. fourcroydes al Cu y sus efectos en el metabolismo de la planta se requieren con el propósito de establecer su nivel de tolerancia para su posible uso en sistemas de fitorremediación como en otras especies de agaves.
CONCLUSIONES
Las concentraciones de cobre añadidas al medio de cultivo no ocasionaron anormalidades fenotípicas en plantas in vitro de henequén var. `Sac Ki' establecidas in vitro ni se observaron síntomas de toxicidad. Se comprobó que el efecto sobre las variables evaluadas se relacionó con la concentración e indicaron tolerancia al cobre. Los resultados de este trabajo sientan las bases para el posible empleo de esta especie en la biorremediación de suelos contaminados. Se sugiere así la realización de más estudios sobre la capacidad fitorremediadora de esta especie junto con su empleo en condiciones de campo ya que los resultados obtenidos la apuntan para tal fin.
REFERENCIAS
Ahmed, A K, Tawfik K M, Zinab A, El-Gawad A (2008) Tolerance of seven faba bean varieties to drought and salt stresses. Research Journal of Agriculture and Biological Sciences 4(2): 175-186
Barceló, J, Poschenrieder, C (2003) Phytoremediation: principles and perspectives. Contributions to Science 2(3): 333-344
Bergmeyer, HU (1974) Methods of Enzymatic Analysis 1. 2nd Edition. Academic Press, New York.
Bradford, MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72: 248-254
Chiarucci, A, Baker, AJM (2007) Advances in the ecology of serpentine soils. Plant and Soil 293: 12
Costa, G, Spitz, E (1997) Influence of cadmium on soluble carbohydrates, free amino acids, protein content of in vitro cultured Lupinus albus. Plant Sci 128: 131-140
Deff, ESH (2007) Cupper treatments and their effects on growth, carbohydrates, minerals and essencial oils contents of Rosmarinu officinalis. World Journal of Agricultural Sciences 3 (3): 322-328
Desikan, R, Hancock, J, Neill, S (2005) Reactive. Blackwell Publishing. Oxford
Küpper, H, Götz, B, Mijovilovich, A, Küpper, Cf, Meyer-Klaucke, W (2009) Complexation and toxicity of copper in higher plants. I. Characterization of copper accumulation, speciation, and toxicity in Crassula helmsii as a new copper accumulator. Plant Physiology 151: 702714
Mahadevan A, Sridhar R (1986) Estimation of total soluble sugar. Methods in physiological plant pathology. 3rd ed. Sivakami Publications: 147-148
Mganga, N, Manoko, MLK, Rulangaranga, ZK (2011) Classification of plants according to their heavy metal content around north mara gold mine, tanzania: implication for phytoremediation. Tanzania Journal of Science 37: 109-119
Mohamed, S I (1986) Growth and yield of tomato and squash in soil treated with Mn. Hort. Sci. 29: 723-730
Nobel, PS (2009) Physicochemical and Environmental Plant Physiology. Fourth Edition. Elsevier. Academic Press. London
Sumner, JB (1921) Dinitrozalicylic acid: a reagent for the estimation of sugar in normal and diabetic. J. Biol. Chem. 47: 5-9
Yruela, I (2005) Copper in plants. Braz. J. Plant Physiol 17(1): 145-156
Recibido: 24-2-2014
Aceptado: 15-4-2014
Copyright (c) 2016 Biotecnología Vegetal
![]()
Biotecnología Vegetal eISSN 2074-8647, RNPS: 2154. ISSN 1609-1841, RNPS: 0397 Editada por: Instituto de Biotecnología de las Plantas. Universidad Central Marta Abreu de Las Villas. Carretera a Camajuaní km 5.5, Santa Clara, Villa Clara, Cuba CP 54 830 Tel: 53 42200124, e-mail: info@ibp.co.cu

Biotecnología Vegetal está bajo una Licencia Creative Commons Atribución-NoComercial 4.0 Internacional.